News
-
2023/1/15
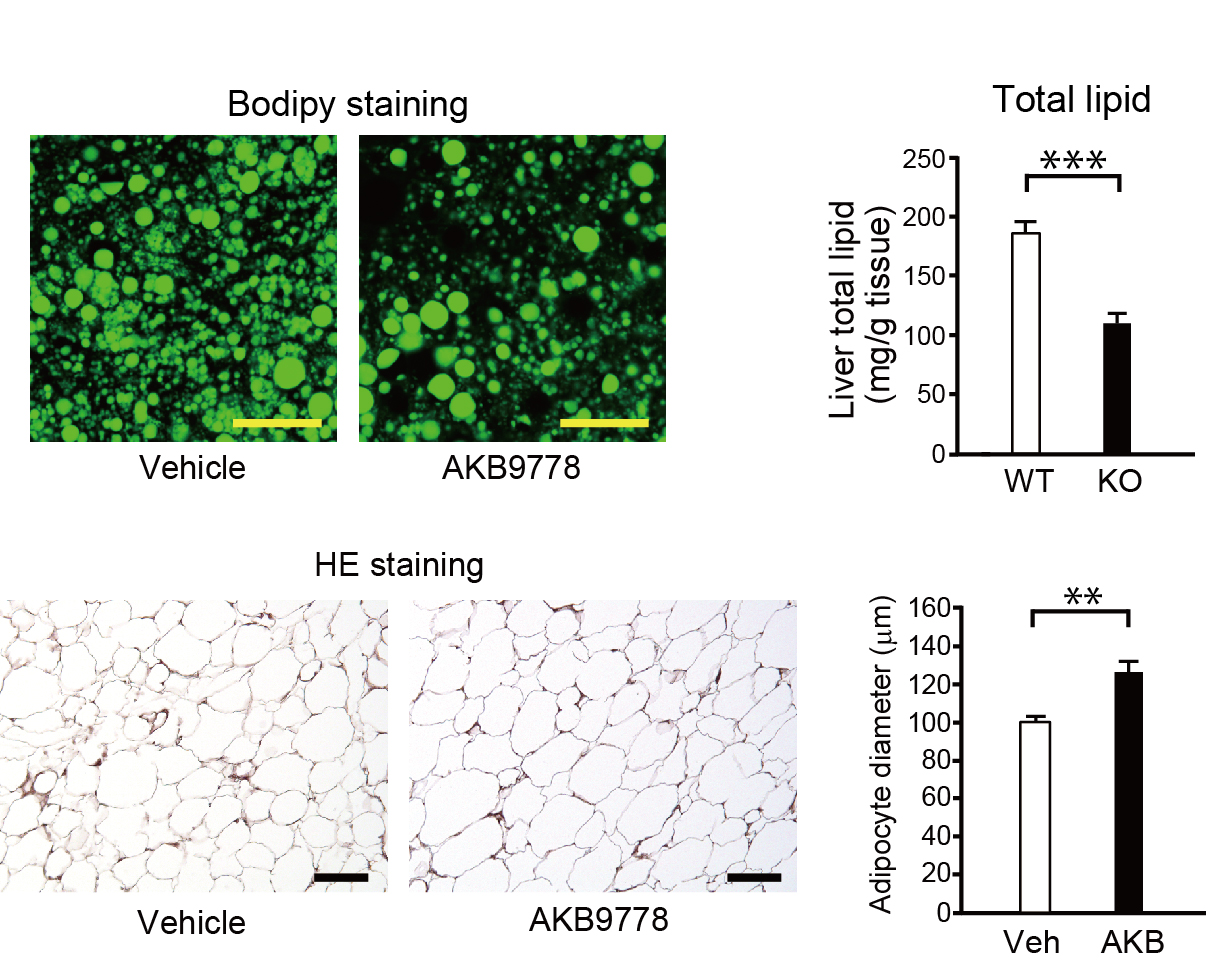
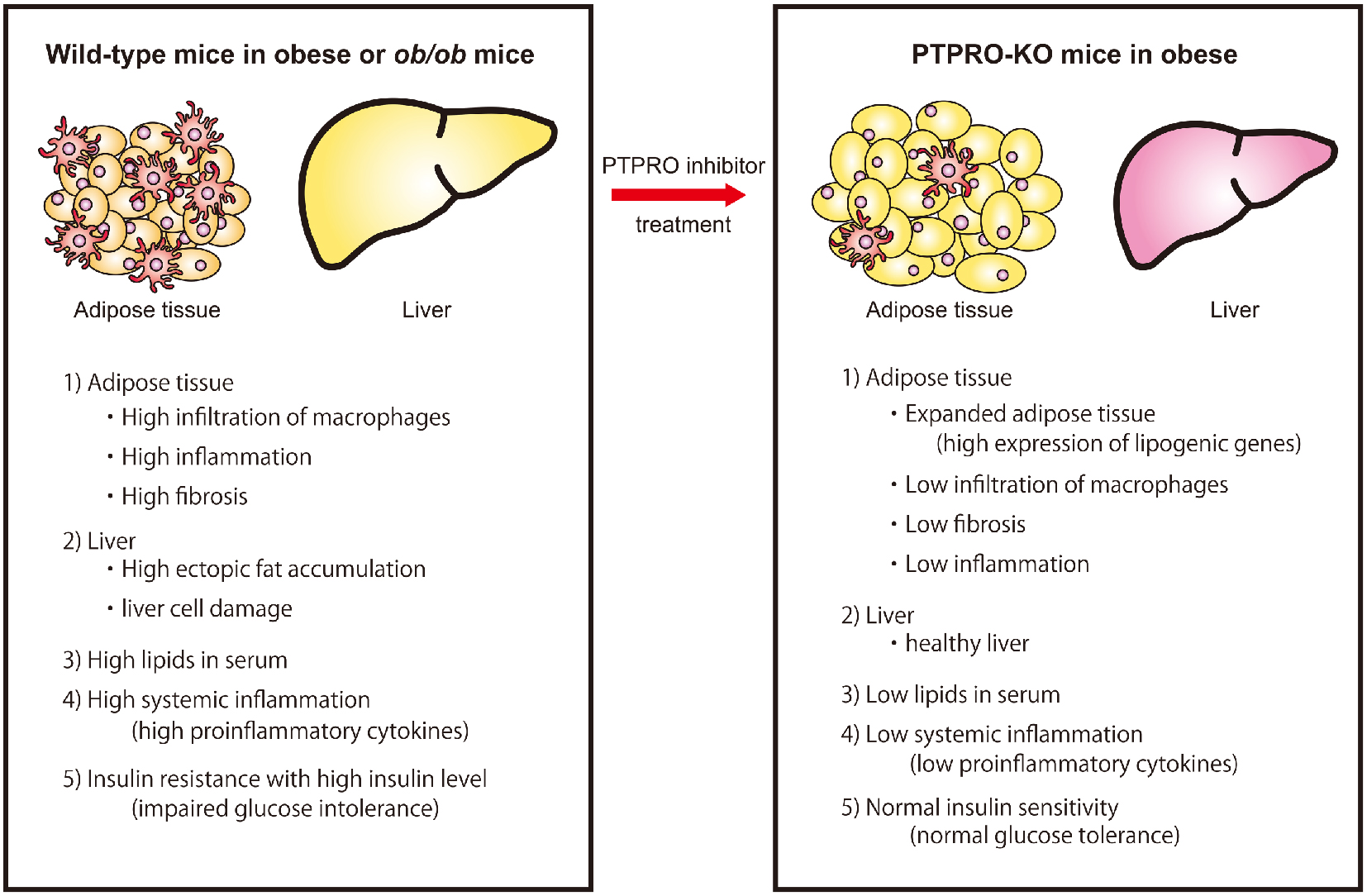
Our paper on “relationship between PTPRO activity and ectopic fat accumulation” was published in Life Sciences.
-
2022/08
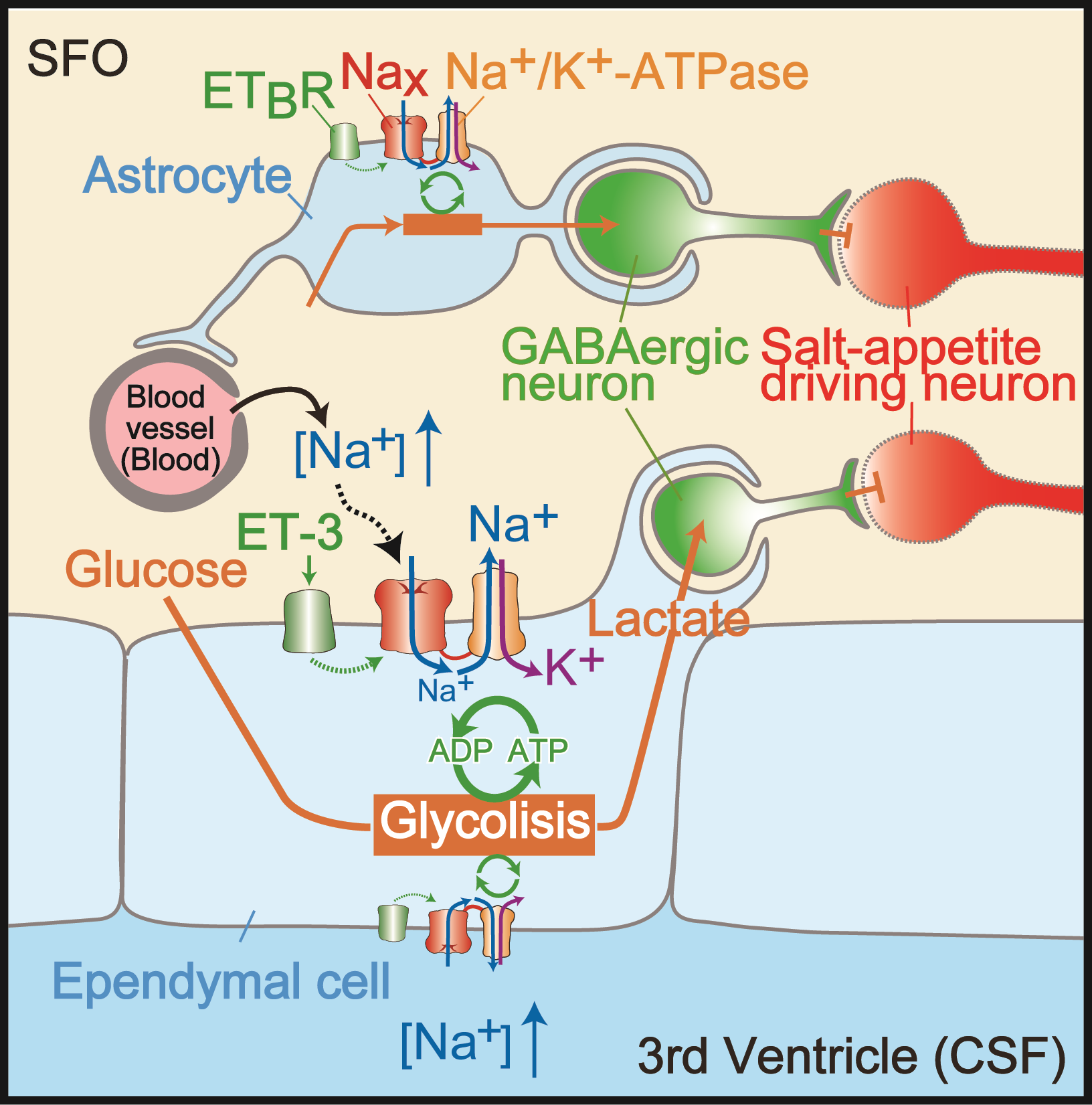
Our review article on “brain mechanisms for water/salt intake control” was published in Proceedings of the Japan Academy, Ser. B, Physical and Biological Sciences.
-
2021/7/29
Professor Dr. Noda won the 23rd Toshihiko Tokizane Memorial Award.
-
2020/11/10
Our research on “water-intake control mechanisms” was published in Nature Communications.
Research
We are currently conducting research aimed at elucidating the intracerebral mechanism of fluid homeostasis and blood pressure control. We are also conducting research on obesity (metabolic control).
I
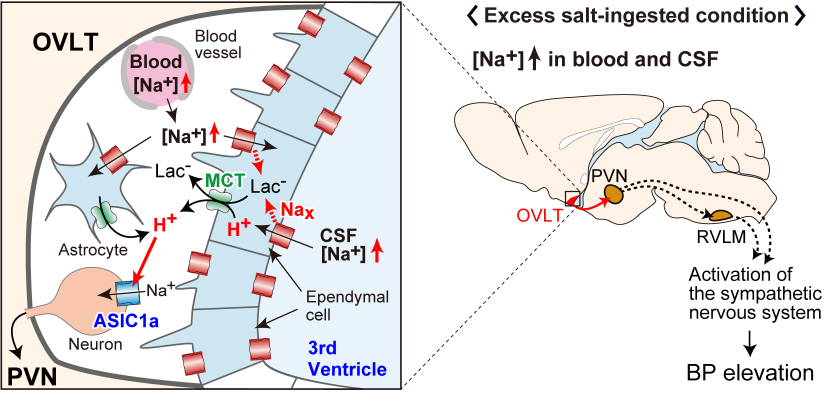
Brain mechanisms controlling blood pressure
We are now promoting the study to elucidate the overall picture of brain mechanisms for ...

II
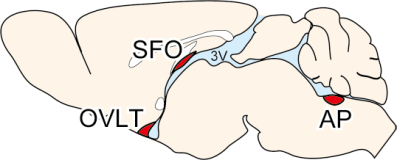
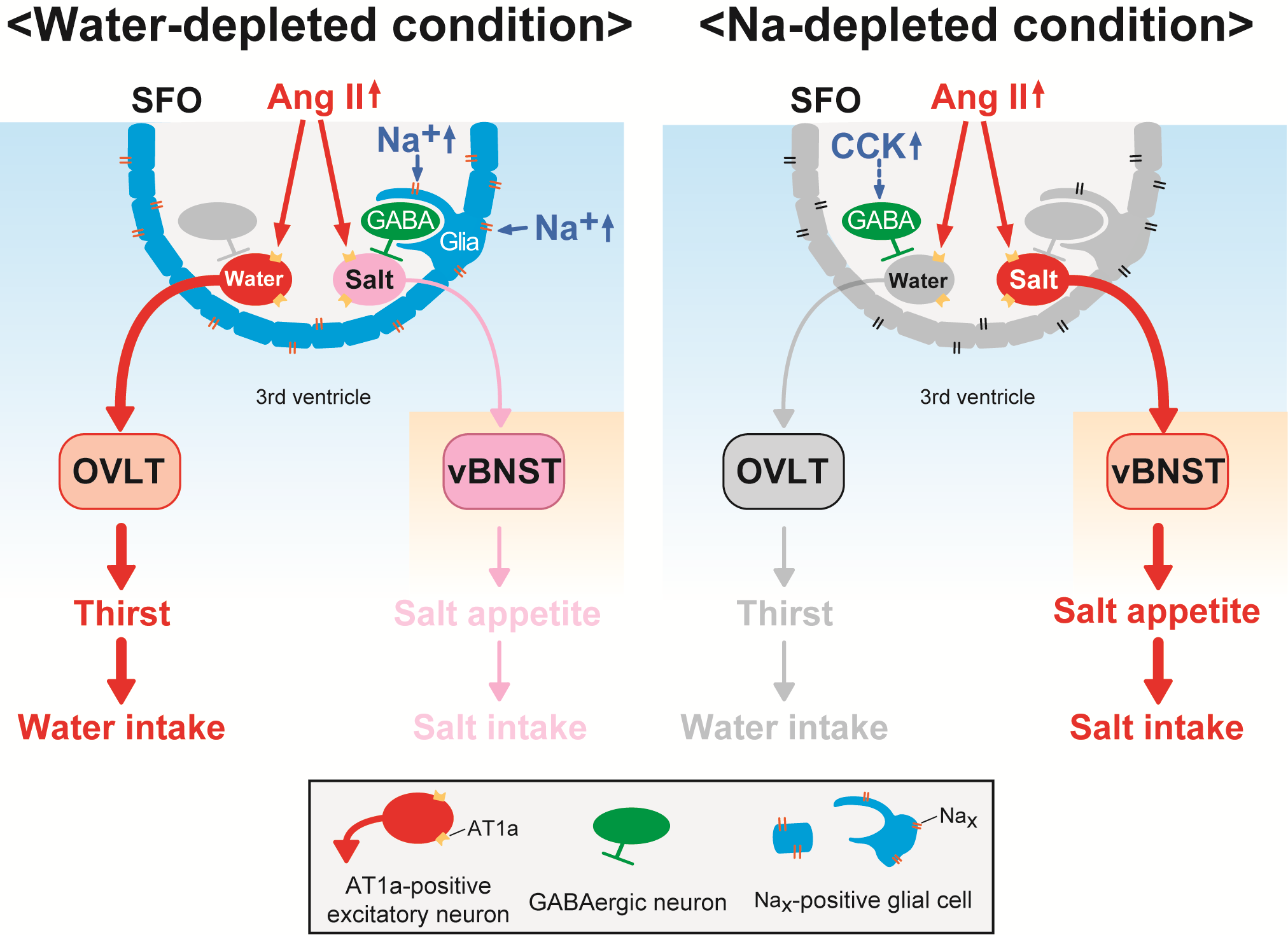
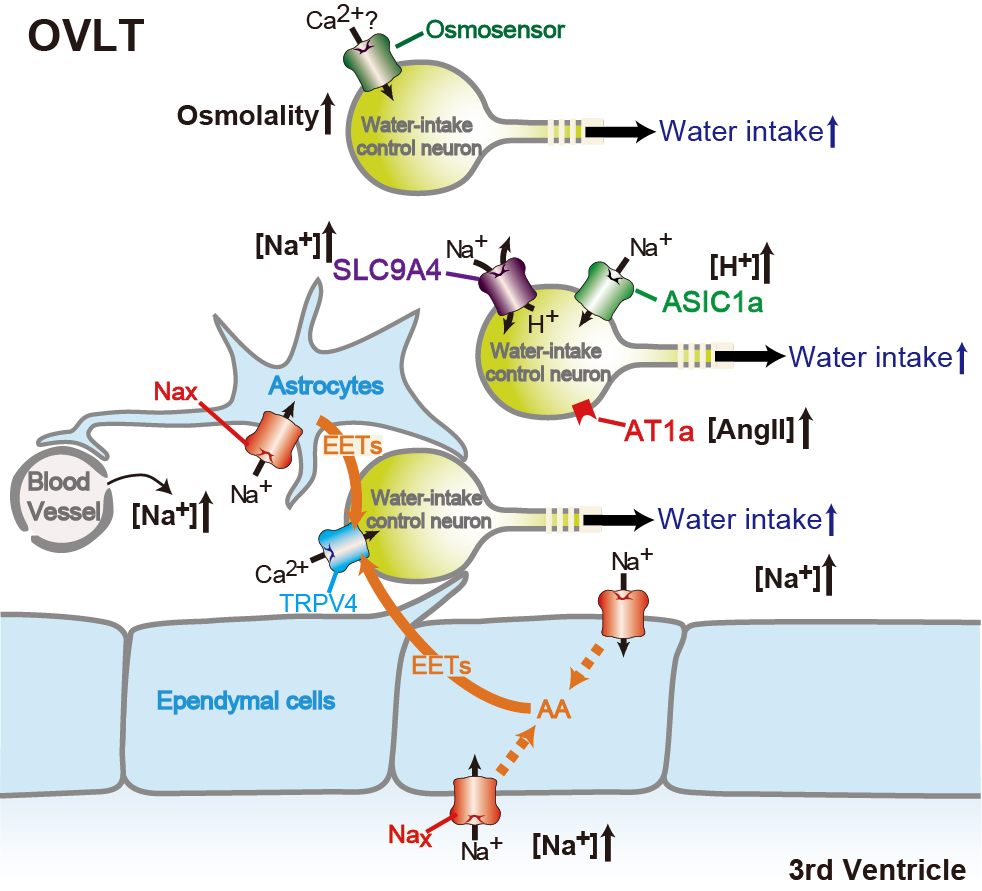
Brain mechanisms controlling thirst and salt appetite
Terrestrial animals, including humans, have mechanisms that maintain body fluid homeostasis ...
III
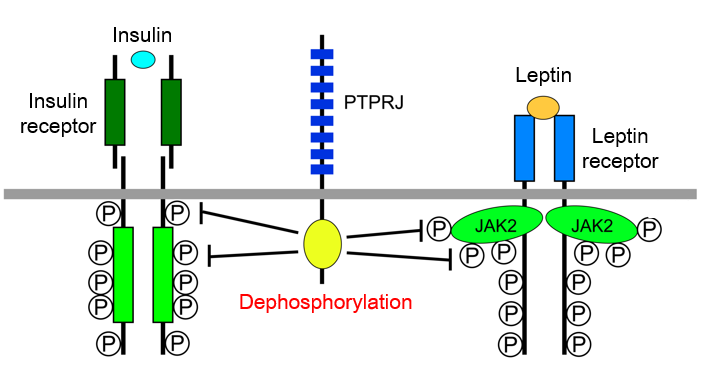
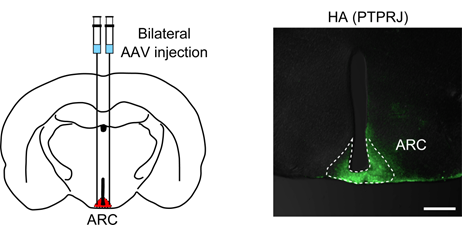
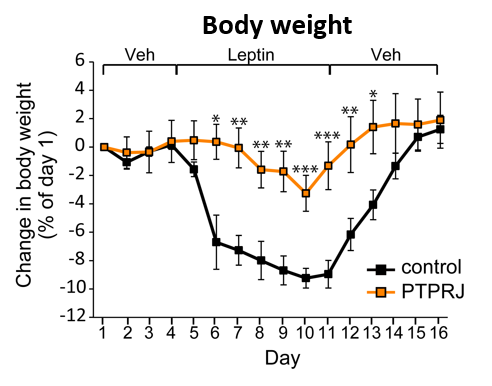
Research on obesity
Obesity is a major risk factor for type 2 diabetes, hypertension, dyslipidemia, and cardiovascular diseases.
People

Masaharu Noda, Ph.D.
Specially Appointed Professor

Takashi Matsuda, Ph.D.
Specially Appointed Associate Professor

Keiko Ikeda, Ph.D.
Visiting Associate Professor (Lecturer)

Wu Siyuan, Ph.D.
Researcher

Yuko Kumeda.
Technical Support Staff
Mami Ishida.
Technical Support Staff
Yutaka Anzai.
Technical Support Staff
Shizuka Miyahara.
Technical Support Staff
Yukari Mutou.
Technical Support Staff
Yuka Shibuya.
Technical Support Staff
Youko Onoda.
Administrative Support Staff


Publications
(Representative recent ones)
-
Nomura K, Hiyama TY, Sakuta H, Matsuda T, Lin CH, Kobayashi K, Kobayashi K, Kuwaki T, Takahashi K,
Matsui S, Noda M.
[Na+] Increases in Body Fluids Sensed by Central Nax Induce Sympathetically Mediated Blood Pressure Elevations via H+-Dependent Activation of ASIC1a.
Neuron 101:60-75. doi: 10.1016/j.neuron.2018.11.017. 2019. -
Matsuda T, Hiyama TY, Niimura F, Matsusaka T, Fukamizu A, Kobayashi K, Kobayashi K, Noda M.
Distinct neural mechanisms for the control of thirst and salt appetite in the subfornical organ.
Nat. Neurosci 2017, 20, 230-241. -
Hiyama TY, Watanabe E, Ono K, Inenaga K, Tamkun MM, Yoshida S, Noda M.
Nax channel involved in CNS sodium-level sensing.
Nature Neurosci. 2002, 5, 511-512.
Photo
Contact
-
Location
4259 Nagatsuta-cho, Midori-ku,
Yokohama, Kanagawa
226-8501 Japan -
Nearest stations
Suzukakedai Campus is a 5-minute walk from Suzukakedai Station on the Tokyu Denentoshi Line. -
Mail
noda.m.19dc@m.isct.ac.jp